Tuesday, 7 August 2012
Tuesday, 31 July 2012

Monday, 30 July 2012
Nervous System of Pila
The central nervous system of the apple snail consists of a several ganglia connected with each other by a commisura. The perikarya (cell bodies of the nerve cells) are concentrated in these ganglia.
Some parts of the nervous system are symmetric (for example the cerebral ganglia) while others (like the subintestinal ganlion) are not, this because of the asymmetric construction of the snail body.
There isn't a central brain in snails like mammals have, the different ganglia are scattered throughout the body of the snail, although mainly located in the head.
Every ganglion enervates some specific organs with its nerves. The pedal ganglia for example enervates the foot (Latin: pedalis) of the snail. The function of the commisures between the ganglia are to integrate the different parts of the nervous system.
Scheme of the nervous system:
Some parts of the nervous system are symmetric (for example the cerebral ganglia) while others (like the subintestinal ganlion) are not, this because of the asymmetric construction of the snail body.
There isn't a central brain in snails like mammals have, the different ganglia are scattered throughout the body of the snail, although mainly located in the head.
Every ganglion enervates some specific organs with its nerves. The pedal ganglia for example enervates the foot (Latin: pedalis) of the snail. The function of the commisures between the ganglia are to integrate the different parts of the nervous system.
Scheme of the nervous system:
 |
Mantle Cavity / Pallial Complex
Mantle cavity:
The mantle cavity is formed by a fold of the backside of the bodywall (roof) and the head neck and body of the snail (floor). It can be considered as a sac, loosely attached to the inside of the shell and enveloping the head-neck and the gills-lung area of the body.
The mantle cavity is formed by a fold of the backside of the bodywall (roof) and the head neck and body of the snail (floor). It can be considered as a sac, loosely attached to the inside of the shell and enveloping the head-neck and the gills-lung area of the body.
 |
The top of the mantle cavity (the part away from the shell opening or the posterior part) is separated in two chambers, the left one forms the lung and right one contains the gill (ctenidium) with the hypobranchial gland in between the rectum and the gill. The latter (hypobranchial gland) secretes mucus into the mantle cavity.
The bottom of the mantle cavity (anterior) is the mantle edge, which fits against the shell opening (aperture) and has an important function: it is the growing edge for the shell.
Besides the gill and the lung, the mantle cavity also contains the penis sheath (in the male snail), female gonopore, rectum, anus and osphradius (smell organ).
The epinaenium is a muscular ridge that starts in the top of the mantle cavity (posterior) and extents across the floor of the mantle (the back of the snail's head) towards the breathing siphon. It's main function is to close the left side were when breating air to prevent an air flow over the gills (right side of the cavity).
The bottom of the mantle cavity (anterior) is the mantle edge, which fits against the shell opening (aperture) and has an important function: it is the growing edge for the shell.
Besides the gill and the lung, the mantle cavity also contains the penis sheath (in the male snail), female gonopore, rectum, anus and osphradius (smell organ).
The epinaenium is a muscular ridge that starts in the top of the mantle cavity (posterior) and extents across the floor of the mantle (the back of the snail's head) towards the breathing siphon. It's main function is to close the left side were when breating air to prevent an air flow over the gills (right side of the cavity).
 The snail body, taken out of the shell (Pomacea diffusa). |  Note: this animal was not killed especially to make this picture. |
 At the front of the foot, there is a cleavage (Pomacea canaliculata). (Click on image to enlarge) |  The muscular foot (Pomaceadiffusa). |
Foot:
The muscular foot is used for locomotion and consists of several muscular layers. There are two types of walking in apple snails: with creeping locomotion and with a small wave locomotion.
The creeping locomotion is consists of an extention-contraction (elongation-shortening) movement of the foot. Snails that use this rather rare technique do not move at a constant speed . At least one apple snail species uses this walking method, although not all the time: Asolene spixi. An example of this can be seen in the following MPEG movie: [Asolene_walk] (140kb). This movie plays at 10x speed. Other species use this movement tecnhique when theu burry in the substrate.
A much more common locomotion is used by the majority of the apple snails and is achived by contraction waves on the sole of the foot. The result is a smooth walk with a constant velocity. An example of this can be seen in this MPEG movie of a Pomacea canaliculata snail: [Pomacea_walk] (210kb). Speed of this movie: 3x.
The muscular foot is used for locomotion and consists of several muscular layers. There are two types of walking in apple snails: with creeping locomotion and with a small wave locomotion.
The creeping locomotion is consists of an extention-contraction (elongation-shortening) movement of the foot. Snails that use this rather rare technique do not move at a constant speed . At least one apple snail species uses this walking method, although not all the time: Asolene spixi. An example of this can be seen in the following MPEG movie: [Asolene_walk] (140kb). This movie plays at 10x speed. Other species use this movement tecnhique when theu burry in the substrate.
A much more common locomotion is used by the majority of the apple snails and is achived by contraction waves on the sole of the foot. The result is a smooth walk with a constant velocity. An example of this can be seen in this MPEG movie of a Pomacea canaliculata snail: [Pomacea_walk] (210kb). Speed of this movie: 3x.
| Movies (MPEG1): - Walking apple snail on glass (Asolene spixi) (140kb, 10x) - Walking Pomacea canaliculata (Pomacea canaliculata) (210kb, 3x) |
Because the apple snail has a lung the animal doesn't have to bear all it's weight when moving under water: the animal partly floats on the air in the lung. This declares why such rather big snails can 'walk' fairly elegant across the bottom.
Besides it locomotion function, the foot is also used to collect food from the surface (film-feeding or ciliary feeding). Forming a funnel with the foot in which particles from the surface can be trapped does this. To attract the floating food pieces, the snail makes the same movement with its foot as it does to walk, but only with the front part of the foot and the middle part, forming the funnel. With the tail of the foot, the snail stays attached to the side or an object near the surface.
Once the funnel is filled with food, the snail brings its mouth in the funnel and starts with its meal.
Besides it locomotion function, the foot is also used to collect food from the surface (film-feeding or ciliary feeding). Forming a funnel with the foot in which particles from the surface can be trapped does this. To attract the floating food pieces, the snail makes the same movement with its foot as it does to walk, but only with the front part of the foot and the middle part, forming the funnel. With the tail of the foot, the snail stays attached to the side or an object near the surface.
Once the funnel is filled with food, the snail brings its mouth in the funnel and starts with its meal.
 Collecting food from the surface, seen from above (Pomaceacanaliculata, juvenile). |  Collecting food from the surface, sideview (Pomacea canaliculata, juvenile). |
Sense Organs
 Apple snails are highly depending on their smell sense. With their smell sense they are able to locate food and recognise other snails of their own species, which is important to find a good mating partner. It's therefore not surprising that apple snail have a well-developed smell sense.
Apple snails are highly depending on their smell sense. With their smell sense they are able to locate food and recognise other snails of their own species, which is important to find a good mating partner. It's therefore not surprising that apple snail have a well-developed smell sense. The vision of the apple snail on the other hand is rather weak (poor ability to form images and bad image quality) and functions merely as a light direction detector.
The tactile sense is well developed as can be seen as the snail walks over a small object or encounters an obstacle.
The hearing capabilities of the apple snail are worthless, even more: they are completely deaf.
Body: The whole body surface of the apple snail contains chemo- and mechanoreceptors.
Eyes: The eyes of the apple snail are located at the base of the tentacles, on top of the eyestalks.
The structure of the eye does not provide detailed vision, they rather function as directional light sensors that give the snail an orientation towards light sources. Colour vision is absent as the retina does not contain colour specific photosensors: an apple snail is colour-blind.
The structure of the eye does not provide detailed vision, they rather function as directional light sensors that give the snail an orientation towards light sources. Colour vision is absent as the retina does not contain colour specific photosensors: an apple snail is colour-blind.
| Movies (MPEG1): - The eyes of an apple snail (Asolene megastoma) (132kb) |
 The eye on the eye stalk of aPomacea canaliculata snail. The eye on the eye stalk of aPomacea canaliculata snail.(click on image to enlarge) | 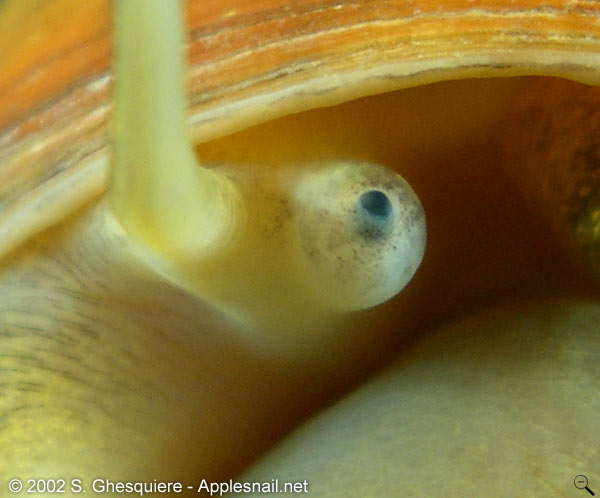 Another photograph of an eye (Pomacea canaliculata). Another photograph of an eye (Pomacea canaliculata).(click on image to enlarge) |
It appears that the eyes of apple snails are rather optimized for optical sensitivity (night vision!) rather than for high detail vision. In fact, two mechanism are used in the apple snail eye to increase sensitivity to light: the eyes are relatively large with a large lens (large eyes= more light captured) and the light sensitive cells (rhabdoms) are grouped and connected with the same neuron (pooling of signals or neural summation = higher sensitivety, but lower resolution). It has even been suggested that the optical sensitivity of the apple snail approaches that of the nocturnal spider Dinopis subrufus (Blest and Land, 1977).
The spatial resolution of the eye, however, is estimated to be around a poor 44 pixels (SEYER, J.O. et al. 1998). This low resolution is due to several factors: the pooled light sensitive cells, the bad quality of the lens (blurring) and unisolated light sensitive cells (high cross-talk between cells). To give an idea, human eyes are capable to resolve 10.000.000 pixels. The pictures below illustrate this even better.
As apple snail do not have colour sensitive cells in their retina, they can only see shades of grey (colour-blind).
One can conclude that the eyes of apple snails are suitable to find light and dark area's within their invironment, even at night, but that their overal vision in the sense of image quality is rather poor.
The spatial resolution of the eye, however, is estimated to be around a poor 44 pixels (SEYER, J.O. et al. 1998). This low resolution is due to several factors: the pooled light sensitive cells, the bad quality of the lens (blurring) and unisolated light sensitive cells (high cross-talk between cells). To give an idea, human eyes are capable to resolve 10.000.000 pixels. The pictures below illustrate this even better.
As apple snail do not have colour sensitive cells in their retina, they can only see shades of grey (colour-blind).
One can conclude that the eyes of apple snails are suitable to find light and dark area's within their invironment, even at night, but that their overal vision in the sense of image quality is rather poor.
 A photograph of a Pomacea diffusashell, cut in half. A photograph of a Pomacea diffusashell, cut in half. |  The same photograph seen through the eyes of an apple snail. Simulated with blur from lens, pooled cells and cross-talk. Image as one would probably receive from the neural cells. |
The anatomy of the apple snail eye is simple and consists of several structures embedded in the connective tissue of the eyestalk: the cornea, the lens, the retina and the optic nerve.
The acellular lens, with a spherical shape, is embedded in the eyestalk. The lens itself consists of a dense ovoid inner core with several peripheral layers at the outside.
The thin cornea on top of the eye forms a circular pupil. This transparant tissue at which light enters the eye consists of 4 layers. The peripheral double layer of the cornea has a cuboidal epithelium layer at the outside and a connective tissue layer with a collagenous structure at the inside. The epithelium at the outside is continuous with the epidermis covering the whole eyestalk and the rest of the body.
The inside of the cornea is part of the lens and is a double layer as well, with microvilli at the inside and an epithelial layer near the periphery.
The retina at the base of the lens is divided in four separate layers. The first layer, surrounding the lens, contains light-sensitive microvillar rhabdoms. The second layer is heavily pigmented and composed of the pigmented parts of the photoreceptor cells and isolated pigment cells. Around this layer is a pigmented layer in which the nuclei of photoreceptor cells are embedded together with supportive cells. The fourth, outermost layer consist of mainly connective tissue containing neural cells and their axons, which connect with the optic nerve.
The acellular lens, with a spherical shape, is embedded in the eyestalk. The lens itself consists of a dense ovoid inner core with several peripheral layers at the outside.
The thin cornea on top of the eye forms a circular pupil. This transparant tissue at which light enters the eye consists of 4 layers. The peripheral double layer of the cornea has a cuboidal epithelium layer at the outside and a connective tissue layer with a collagenous structure at the inside. The epithelium at the outside is continuous with the epidermis covering the whole eyestalk and the rest of the body.
The inside of the cornea is part of the lens and is a double layer as well, with microvilli at the inside and an epithelial layer near the periphery.
The retina at the base of the lens is divided in four separate layers. The first layer, surrounding the lens, contains light-sensitive microvillar rhabdoms. The second layer is heavily pigmented and composed of the pigmented parts of the photoreceptor cells and isolated pigment cells. Around this layer is a pigmented layer in which the nuclei of photoreceptor cells are embedded together with supportive cells. The fourth, outermost layer consist of mainly connective tissue containing neural cells and their axons, which connect with the optic nerve.
 Overview illustration of the eye. |  Close up of the cornea. |
Tentacles: The tentacles are very important sensory organs. Apple snails highly rely on the smell capacity and the sensitivity of their tentacles to navigate in their environment.
The cephalic tentacles (attached at the top of the head) can be very long, sometimes even longer than the body of the snail.
The labial tentacles or labial palps (attached at each side of the mouth), a typical feature of apple snails, are shorter.
Osphradia: The osphradius is a chemosensory structure that is located in the mantle cavity, in front of the lung.
The osphradius gives the snail the capability to smell chemical substances in the water.
The cephalic tentacles (attached at the top of the head) can be very long, sometimes even longer than the body of the snail.
The labial tentacles or labial palps (attached at each side of the mouth), a typical feature of apple snails, are shorter.
Osphradia: The osphradius is a chemosensory structure that is located in the mantle cavity, in front of the lung.
The osphradius gives the snail the capability to smell chemical substances in the water.
 |  Osphradia near the opening of thelung (Pomacea diffusa). |
The statocysts: The statocysts (2) are vesicles containing a statolith (little stone like structures composed of calcium carbonate). They function as balancing organs, used by the snail to detect its position with regard to the ground.
They are located inside the snail's body close to the pedal ganglia.
They are located inside the snail's body close to the pedal ganglia.
Reproductive System of Pila
The apple snail has separated sexes (dioecious) and is NOT a hermaphroditic snail.
The differences between the sexes are not visible at the shell colour or patterns, but in several species, a difference in shell shape has been described. For example in Pomacea canaliculata, the aperture (opening) in the male has a rounder opening than in the female because of the large penial complex. This sex-difference is not very well visible and it's not a very reliable to dicriminate on individual base, but clear when measuring large amount of snails (in other words, it's possible to show the difference based on statistics).
In some species however, the difference between male and female are very obvious in the adult snails. For example in Pomacea flagellata flagellata, the male has a very wide shell lip (the shell opening actually has the shape of a trumpet) when reaching sexual maturity. This hasn't been described in the literature yet (october 2000), but the author of this site has observed this in his entire Flagellata population. This shell shape difference cannot be explained by the space occupied by the penial complex, but is rather a reproduction strategy: the male can block the entrance of the female's mantle cavity during mating, in order to prevent other males from copulating with the female. Note that this is only a hypothesis of the author. No serious scientific research has been carried out on this topic.
The differences between the sexes are not visible at the shell colour or patterns, but in several species, a difference in shell shape has been described. For example in Pomacea canaliculata, the aperture (opening) in the male has a rounder opening than in the female because of the large penial complex. This sex-difference is not very well visible and it's not a very reliable to dicriminate on individual base, but clear when measuring large amount of snails (in other words, it's possible to show the difference based on statistics).
In some species however, the difference between male and female are very obvious in the adult snails. For example in Pomacea flagellata flagellata, the male has a very wide shell lip (the shell opening actually has the shape of a trumpet) when reaching sexual maturity. This hasn't been described in the literature yet (october 2000), but the author of this site has observed this in his entire Flagellata population. This shell shape difference cannot be explained by the space occupied by the penial complex, but is rather a reproduction strategy: the male can block the entrance of the female's mantle cavity during mating, in order to prevent other males from copulating with the female. Note that this is only a hypothesis of the author. No serious scientific research has been carried out on this topic.

Despite the shape difference, it often easier to determine the sex by looking inside the mantle cavity to spot the penial complex or the absence thereof in case of a female snail. To do this one should take the snail out of the water and keep it upside down for a minute or 10 and blow some fresh air over the snails body. The animal will (hopefully) stretch its foot out to turn the shell upright. When this happens it's possible to look inside the right mantle cavity, where one should look for the relatively large penissheath in front of the gills. The female snail lack such structure (obvious). The picture above should help to see the difference between the sexes.
Last, but not least: in very thin shelled apple snails and in specimen with translucent shell (albino Pomacea diffusa), one can also observe a colour difference in the upper whorl (spine) of the shell in periods that the snails are in a reproduction stage. The female ovaria can then show up as a dark organ at that place (see picture below). Note however, that this method is not reliable at all times and can only be used when the snail are sexual active. In the latter case the snails mate anyway, so that one can see the male snail mounting the female repeatedly.
A sex change from male to female (proandry) has been described for some Pila and Pomacea apple snails. In case of Pila snails, the male needs a preparatory aestivation period to become a female, while thePomacea snails can change sex at all times (at least in certain species). During this sex change, the testis change in colour and decrease in size. The frequency at which this sex change occurs is not certain and not much literature is available about this subject. There is some very interesting research about the effects of certain chemical substances that are known of suspected to have influence on the development of the reproductive organs. Many of these substances are of great concern as they can be found in habitats due to pollution. Check here for an interesting research page about this subject (in German).
Last, but not least: in very thin shelled apple snails and in specimen with translucent shell (albino Pomacea diffusa), one can also observe a colour difference in the upper whorl (spine) of the shell in periods that the snails are in a reproduction stage. The female ovaria can then show up as a dark organ at that place (see picture below). Note however, that this method is not reliable at all times and can only be used when the snail are sexual active. In the latter case the snails mate anyway, so that one can see the male snail mounting the female repeatedly.
A sex change from male to female (proandry) has been described for some Pila and Pomacea apple snails. In case of Pila snails, the male needs a preparatory aestivation period to become a female, while thePomacea snails can change sex at all times (at least in certain species). During this sex change, the testis change in colour and decrease in size. The frequency at which this sex change occurs is not certain and not much literature is available about this subject. There is some very interesting research about the effects of certain chemical substances that are known of suspected to have influence on the development of the reproductive organs. Many of these substances are of great concern as they can be found in habitats due to pollution. Check here for an interesting research page about this subject (in German).
 When snails with a tranlucent shell (albino Pomacea diffusa in this case) are ready to reproduce, the ovaria of the female (left) can become suspicious through the shell. |  Marisa cornuarietis is not selective when it comes to a mating partner and mounts other species as well (in this case a Pomacea flagellatasnail). |  Sex differences in adult Pomaceaflagellata flagellata snails. The female is larger and has a straight shell opening, while the male (left) has a extended shell lip. |
Sex ratio
The sex ratio within apple snail populations is around 0.5, which means that there is an equal amount of males and females. However, the sex ratio between the broods of a population varies a lot. Some egg masses give almost exclusively male hatchlings, while other give exclusively female offspring. It was first thought that environmental factors influence the sex ratio in apple snails. For example the temperature could determine of the snail become male or female, like is the case for crocodiles.
Recent research (Yusa & Suzuki, 2003) shows that the sex ratio is likely genetical determined. It turns out that different spawns of a single mating pair shows little variation in sex ratio between the different broods, despite variations in environmental circumstances. This supports a genetic basis for sex determination.
In humans, the sex of an individual is determined by the X and the Y chromosomes (XX= female, XY= male). However, there is no clear evidence for the presence of X and Y chromosomes in apple snails. Most likely, apple snails have a different sex determining system, in which sex is determined by a number of genes (polyfactorial sex determination). The combination of at least three and most likely more genes determine the sex ratio. Future research will hopefully give a better insight in the sex determining mechanisms.
The sex ratio within apple snail populations is around 0.5, which means that there is an equal amount of males and females. However, the sex ratio between the broods of a population varies a lot. Some egg masses give almost exclusively male hatchlings, while other give exclusively female offspring. It was first thought that environmental factors influence the sex ratio in apple snails. For example the temperature could determine of the snail become male or female, like is the case for crocodiles.
Recent research (Yusa & Suzuki, 2003) shows that the sex ratio is likely genetical determined. It turns out that different spawns of a single mating pair shows little variation in sex ratio between the different broods, despite variations in environmental circumstances. This supports a genetic basis for sex determination.
In humans, the sex of an individual is determined by the X and the Y chromosomes (XX= female, XY= male). However, there is no clear evidence for the presence of X and Y chromosomes in apple snails. Most likely, apple snails have a different sex determining system, in which sex is determined by a number of genes (polyfactorial sex determination). The combination of at least three and most likely more genes determine the sex ratio. Future research will hopefully give a better insight in the sex determining mechanisms.
 |
Male reproductive system
The male reproductive tract consist of the testis and vas deferens, the seminal vesicle and the prostate gland, the penial sac, the penis and the penis sheath.
The cream-yellow testis can be found in the upper part (1.5 to 2.5 coils) of the shell along the dark coloured digestive gland. The short tubules (vasa efferentia) inside the testis fuse to form the vas deferens.
The vas deferens passes down to the seminal vesicle, beneath the kidney chamber. In this seminal vesicle, the sperm cells are stored.
The prostate gland is closely connected with the seminal vesicle and is situated next to the seminal vesicle (they appear as one structure from the outside). The prostate gland passes down the right margin of the mantle skirt, along the rectum and ends in the penial papilla, next to the anal papilla at the roof of the rightmantle cavity.
During mating activities, this penial papilla bends towards the sperm groove in which the sperm is conducted. From this sperm groove the sperm is conveyed in the penial duct at the base of the penis.
The penis itself is coiled in a basal pouch (penial sac) when not used. On erection the penis comes out of the penial sac and is grasped around on the lower thirth by a muscular penial sheath from the mantle. It's the latter, which can be seen when the snails are mating. The real penis is rather thin, whip-like and often stays out of sight.
The male reproductive tract consist of the testis and vas deferens, the seminal vesicle and the prostate gland, the penial sac, the penis and the penis sheath.
The cream-yellow testis can be found in the upper part (1.5 to 2.5 coils) of the shell along the dark coloured digestive gland. The short tubules (vasa efferentia) inside the testis fuse to form the vas deferens.
The vas deferens passes down to the seminal vesicle, beneath the kidney chamber. In this seminal vesicle, the sperm cells are stored.
The prostate gland is closely connected with the seminal vesicle and is situated next to the seminal vesicle (they appear as one structure from the outside). The prostate gland passes down the right margin of the mantle skirt, along the rectum and ends in the penial papilla, next to the anal papilla at the roof of the rightmantle cavity.
During mating activities, this penial papilla bends towards the sperm groove in which the sperm is conducted. From this sperm groove the sperm is conveyed in the penial duct at the base of the penis.
The penis itself is coiled in a basal pouch (penial sac) when not used. On erection the penis comes out of the penial sac and is grasped around on the lower thirth by a muscular penial sheath from the mantle. It's the latter, which can be seen when the snails are mating. The real penis is rather thin, whip-like and often stays out of sight.
|



In the sperm cell production in Pomacea canaliculata and perhaps also all other members of the Ampullariidae family, two types of sperms are produced: eupyrene sperm, or normal, viable sperms cells, and apyrene parasperm. This latter type of sperm consists of sterile cells, which are the result of a complex cytodifferentiation process, in which they develop into anuclear ciliated cells, completely different from the normal sperm cells. The apyrene paraspermatozoa (sperm cells) are filled with secretory granules, rich in glycoproteins (an energy rich protein molecule).
The function of these sterile sperms cells is not fully understood, but several possible functions are proposed. In one hypothesis, the package of normal sperm cells (euspermatozoa) with these sterile (apyrene paraspermatozoa) cells into large motile units, is considered as a solution to penetrate the sperm plug left inside the female to block access for sperms of concurrent males.
Providing nutrition for the functional (eupyrene) sperm cells, could be another function of the apyrene sperm cells. According to this viewpoint, the apyrene sperm cells support the normal sperm cell in their travel trough the female reproductive tract by releasing their glycoproteins. These glycoproteins are an important energy source for the moving sperm cells.
The function of these sterile sperms cells is not fully understood, but several possible functions are proposed. In one hypothesis, the package of normal sperm cells (euspermatozoa) with these sterile (apyrene paraspermatozoa) cells into large motile units, is considered as a solution to penetrate the sperm plug left inside the female to block access for sperms of concurrent males.
Providing nutrition for the functional (eupyrene) sperm cells, could be another function of the apyrene sperm cells. According to this viewpoint, the apyrene sperm cells support the normal sperm cell in their travel trough the female reproductive tract by releasing their glycoproteins. These glycoproteins are an important energy source for the moving sperm cells.
The production of an egg in Pomacea: |
Female reproductive system
The egg cells of the female snail are stored in the ovaria, which are located in the top of the spine, closely to the digestive gland.
To produce an egg, the egg cell is brought to the receptaculum seminis. In this place, where the sperm from the male is stored, the fertilisation of the egg cells takes place. The spermatozoa (sperm cells) of the male can survive for more than a month in the female, and several egg clutches can be fertilised with a single copulation.
The albumen (yolk) from which the snail embryo is living is added by the proximal albumen gland and further during its way through the eggs tube by the distal albumen gland. These glands increase considerably when the female is about to produce eggs. And while the ovaria are located in the top of the shell, the albumen glands are located in the top of the mantle cavity and in the reproductional season they move closer to the shell opening and can even fill a large part of the right mantle cavity.
The egg cells of the female snail are stored in the ovaria, which are located in the top of the spine, closely to the digestive gland.
To produce an egg, the egg cell is brought to the receptaculum seminis. In this place, where the sperm from the male is stored, the fertilisation of the egg cells takes place. The spermatozoa (sperm cells) of the male can survive for more than a month in the female, and several egg clutches can be fertilised with a single copulation.
The albumen (yolk) from which the snail embryo is living is added by the proximal albumen gland and further during its way through the eggs tube by the distal albumen gland. These glands increase considerably when the female is about to produce eggs. And while the ovaria are located in the top of the shell, the albumen glands are located in the top of the mantle cavity and in the reproductional season they move closer to the shell opening and can even fill a large part of the right mantle cavity.
| Microscopic images: - detail of the albumen gland (Pomacea canaliculata) (64k) |
On its way to the shell gland, where the calcareous outside of the egg is added (for the species that lay their eggs above the waterline), the egg is passes through the pseudo-bursa copulatrix. In this bursa the sperms of the male are received and the defect or incomplete sperm cell that aren't able to swim towards the receptaculum seminis, are absorbed here.
At the end of the egg tube, the egg is finished and leaves the genital tract at the gonopore.
At the end of the egg tube, the egg is finished and leaves the genital tract at the gonopore.
| Movies (MPEG1): - Egg deposition (Pomacea diffusa) (267kb) |
The apple snail species from the genera Pila and Pomacea deposit their eggs above the waterline. They are guided to the egg clutch in an ovipositor groove over the snail's body. This groove lays on top of a swollen part of the foot, which is formed by inflating the tissue below with fluid. The ovipositor structure is, as result of this, visible as a semi-transparant lump on the snails body. Note however, that not all apple snails exhibit such visible structure when depositing their eggs. One of the species which has a good visible ovipositor structure is Pomacea diffusa. To get a better idea about the eggs deposition, watch this mpeg movie of a Pomacea diffusa snail depositing her eggs: [egg_laying_movie] (267kb). Note that the speed of the movie has been increased (25 times real time).
The eggs of the species that deposit their eggs above the waterline are soft when they appear from the snails body, but the eggshell hardens within a few hours after they are laid.
The eggs of the species that deposit their eggs above the waterline are soft when they appear from the snails body, but the eggshell hardens within a few hours after they are laid.
| Interactive 3D-models (Java): - Aquatic eggs on a stem (95kb) |
However not all apple snail lay their eggs above the waterline: the snails from the genera Asolene, Lanistes, Marisa and possibly also Afropomus and Saulea (anyone info about this?) lay their eggs in the water, embedded in a gelatinous mass. These eggs are relatively small when they are laid, but take up a lot of water within the first hours after being laid and thus swell significant.
The rate of which the eggs are laid is about 1 every 30 seconds in Pomacea canaliculata and Pomacea diffusa.
The rate of which the eggs are laid is about 1 every 30 seconds in Pomacea canaliculata and Pomacea diffusa.
 Snail depositing her eggs above the waterline (Pomaceacanaliculata). |  Eggs, Pomacea canaliculata, 7 days old. |  The bright green eggs of Pomaceahaustrum. (picture not licenced under creative commons) |
Circulatory System Of Pila


{kind=link}
The blood circulation of the apple snail is a typical example of the circulation in a monocardia: there is only one auricle that receives oxygen rich blood from the lung and the gills and deoxygenated blood from the kidney. So there is no separated blood circulation for oxygen rich and deoxygenated blood like in mammals and birds. It's a less efficient system, but it fulfils the needs of a snail very well.
The blood of apple snails (and snails in general) has two functions: transport of O2, CO2, hormones, nutrition and waste products and a structural function: a hydroskeleton.
The transport capacity of the blood for O2 and CO2 is enhanced by the chemical substance hemocyanine in the blood cells. Hemocyanine fulfils the same function as haemoglobin does in mammals (binding O2 and CO2 to ease transportation), but is colourless in contrary to the red colour of haemoglobin.
As the body of a snail does not contain a skeleton to support the extension movements, for example stretching out a tentacle, snails have to use another way: regulating the blood pressure in the body parts. In other words: inflating and deflating parts of the body in combination of muscle contraction to change shape. The regulation of the local blood is obtained by controlling the input and output of the bloodflow by contracting and relaxing small muscles that surround the veins.
Retracting movements are done by simple muscle contraction, without the need of fluid transportation.
The transport capacity of the blood for O2 and CO2 is enhanced by the chemical substance hemocyanine in the blood cells. Hemocyanine fulfils the same function as haemoglobin does in mammals (binding O2 and CO2 to ease transportation), but is colourless in contrary to the red colour of haemoglobin.
As the body of a snail does not contain a skeleton to support the extension movements, for example stretching out a tentacle, snails have to use another way: regulating the blood pressure in the body parts. In other words: inflating and deflating parts of the body in combination of muscle contraction to change shape. The regulation of the local blood is obtained by controlling the input and output of the bloodflow by contracting and relaxing small muscles that surround the veins.
Retracting movements are done by simple muscle contraction, without the need of fluid transportation.
| Movies (MPEG1): - Heartbeat seen through shell (Pomacea canaliculata) (97kb) |
| Snail heart in action: | |
 |  |
| Pomacea canaliculata. | |
The transport of the blood to and from the organs occurs through arteries (from heart to organs) and veins (from organs to heart). Snails don't have capillary veins and arterioles, which means their blood doesn't flow within tube-like structures (veins and arteries) during the whole circulation, but at the tissue level the blood circulates free between the cells and structures embedded in blood cavities (hemocoels) within the body (=open circulation).
The circulation and filtration of the blood:
The heart of apple snails is well developed and consists of two chambers: the auricle and the ventricle.
The auricle is which receives the blood influx from the lung and the kidney veins is much smaller then the ventricle. Inside the auricle, there are many small muscle fibres connecting the opposite wall trough the lumen. The walls have a spongy surface at the inside and a basal layer at the outside. The blood is able to reach the basal membrane through the spongy surface. Contraction is achieved by contracting these muscle fibres.
The ventricle is much larger and has thick, muscular walls with many spaces between the wall muscles, allowing the blood to be trapped in these semi-vescicles and filtered through the basal membrane. In contrast with the auricular contraction, the ventricular contraction is based on contraction of the wall muscles. The mean ventricular pulse pressure of the African apple snail Lanistes carinatus is reported to be around 7.8 cm of water.
The aortic ampulla functions as a compensation sac and compensates the elevated blood pressure in the aorta during the contraction of the ventricle. Besides its function to regulate the blood pressure, the ampulla also has a function in the immune system as the walls of the ampulla consists of vacuolated tissue with many phagocytes in it. These phagocytes possibly eliminate micro-organisms from the bloodflow. The wall of the ampulla is relatively impermeable, excluding the ampulla for blood filtration.
Both the heart and the ampulla are embedded in the pericard, which is connected with the posterior kidney through a renopericadial canal. The walls of the pericard cover the heart and the ampulla and in the pericard cavity the walls are covered with microvilli, small intercellular channels and ridges. Near the renopericardial canal there are some mucous cells secreting mucus, presumably to bind small particles to be transported to the kidney.
The fluid excreted in the pericard cavity can be considered to be primary urine and this fluid is transported to the kidneys for further filtration and resorption of usable compounds (sodium and chlorine).
The kidney or nephridium consists of two parts: the posterior chamber which excretes uric acid and purines and the anterior chamber which has osmoregulatory function.
The heart of apple snails is well developed and consists of two chambers: the auricle and the ventricle.
The auricle is which receives the blood influx from the lung and the kidney veins is much smaller then the ventricle. Inside the auricle, there are many small muscle fibres connecting the opposite wall trough the lumen. The walls have a spongy surface at the inside and a basal layer at the outside. The blood is able to reach the basal membrane through the spongy surface. Contraction is achieved by contracting these muscle fibres.
The ventricle is much larger and has thick, muscular walls with many spaces between the wall muscles, allowing the blood to be trapped in these semi-vescicles and filtered through the basal membrane. In contrast with the auricular contraction, the ventricular contraction is based on contraction of the wall muscles. The mean ventricular pulse pressure of the African apple snail Lanistes carinatus is reported to be around 7.8 cm of water.
The aortic ampulla functions as a compensation sac and compensates the elevated blood pressure in the aorta during the contraction of the ventricle. Besides its function to regulate the blood pressure, the ampulla also has a function in the immune system as the walls of the ampulla consists of vacuolated tissue with many phagocytes in it. These phagocytes possibly eliminate micro-organisms from the bloodflow. The wall of the ampulla is relatively impermeable, excluding the ampulla for blood filtration.
Both the heart and the ampulla are embedded in the pericard, which is connected with the posterior kidney through a renopericadial canal. The walls of the pericard cover the heart and the ampulla and in the pericard cavity the walls are covered with microvilli, small intercellular channels and ridges. Near the renopericardial canal there are some mucous cells secreting mucus, presumably to bind small particles to be transported to the kidney.
The fluid excreted in the pericard cavity can be considered to be primary urine and this fluid is transported to the kidneys for further filtration and resorption of usable compounds (sodium and chlorine).
The kidney or nephridium consists of two parts: the posterior chamber which excretes uric acid and purines and the anterior chamber which has osmoregulatory function.
 MRI image of a male apple snail (Pomacea diffusa). |
The heart action:
 Au = Auricle V = Ventricle |
The posterior kidney chamber receives the primary urine from the pericard cavity. The folds on the posterior chamber wall have a dense vascular network and are covered with excretory, ciliated and mucous cells. The excretory cells excrete uric acids and other purines from the blood into the lumen of the chamber in which the primary urine flows.
The anterior kidney chamber differs from the posterior chamber in that the lumen is occluded with large lamina that covers the walls. These lamina remarkably increase the surface area that comes in contact with the urine. The epithelium on these lamina is almost entirely consisting of resorptive cells that presumable resorb ions from the urine into the blood.
The renal aperture (urine opening) is situated in the upper region of the right mantle cavity. The urine produced by the kidneys is expelled here.
The anterior kidney chamber differs from the posterior chamber in that the lumen is occluded with large lamina that covers the walls. These lamina remarkably increase the surface area that comes in contact with the urine. The epithelium on these lamina is almost entirely consisting of resorptive cells that presumable resorb ions from the urine into the blood.
The renal aperture (urine opening) is situated in the upper region of the right mantle cavity. The urine produced by the kidneys is expelled here.
The aorta with it's white calcareous granula in it's wall consists two parts: the anterior and the posterior part.
The anterior aorta connects the heart with the head (cephalic hemocoel) and the foot (foot hemocoel), while the posterior aorta divides close to the heart with one artery distributing the blood to the digestive system and the second serves several other organs (testis, ovaria, intestines etc.).
After circulating through the tissues and hemocoels of the snail, the blood is collected in large veins and brought to the posterior kidney.
A portion of the blood that enters the posterior kidney directly flows back to the heart, while the remaining blood enters the vascular system of the kidneys (posterior and anterior part).
After passing the anterior kidney, the blood flows through the mantle vein, from where many small veins bring the blood to the gills and the lung, where O2 uptake and CO2 is exchange takes place.
The lung-gill vein collects the oxygen rich blood from the lung and the gills and brings it back to the heart.
The anterior aorta connects the heart with the head (cephalic hemocoel) and the foot (foot hemocoel), while the posterior aorta divides close to the heart with one artery distributing the blood to the digestive system and the second serves several other organs (testis, ovaria, intestines etc.).
After circulating through the tissues and hemocoels of the snail, the blood is collected in large veins and brought to the posterior kidney.
A portion of the blood that enters the posterior kidney directly flows back to the heart, while the remaining blood enters the vascular system of the kidneys (posterior and anterior part).
After passing the anterior kidney, the blood flows through the mantle vein, from where many small veins bring the blood to the gills and the lung, where O2 uptake and CO2 is exchange takes place.
The lung-gill vein collects the oxygen rich blood from the lung and the gills and brings it back to the heart.
Respiratory System of Pila
The respiratory system of the apple snail is highly adapted to tropical waters, which once in a while dry up. This change in water availability is reflected in the apple snails respiration system: a combination of both a gill and a lung.
The arise of the lung in apple snail can be explained by the following conditions:
Low oxygen levels:
The oxygen (O2) levels in tropical pools can become very low due to the combination of high temperatures and a lot of rotting organic waste. Aerial respiration is a efficient way to overcome this lack of oxygen in the water and is not only used by snails, but also certain fishes have developed an organ to derive oxygen from air (for example labyrinth fishes like Betta splendens and others).
Periods of drought:
Many pools and small rivers in tropical regions dry up during periods of drought. In these periods apple snails aestivate buried in the mud and having only a gill would drastically decrease the chance to survive in such condition.
Availability of food:
Having a lung increases the action radius of the snail. On other word: the snails can leave the water for short periods to gather food above the water surface.
The arise of the lung in apple snail can be explained by the following conditions:
Low oxygen levels:
The oxygen (O2) levels in tropical pools can become very low due to the combination of high temperatures and a lot of rotting organic waste. Aerial respiration is a efficient way to overcome this lack of oxygen in the water and is not only used by snails, but also certain fishes have developed an organ to derive oxygen from air (for example labyrinth fishes like Betta splendens and others).
Periods of drought:
Many pools and small rivers in tropical regions dry up during periods of drought. In these periods apple snails aestivate buried in the mud and having only a gill would drastically decrease the chance to survive in such condition.
Availability of food:
Having a lung increases the action radius of the snail. On other word: the snails can leave the water for short periods to gather food above the water surface.
Although apple snails do have developed a lung, they retained their gill during the course of evolution, this in contrary to the Pulmonata snails, which only have a lung and no gill.
Having a gill provides a big advantage, as the snail does not have to come to the surface that often. At the surface the apple snail is more vulnerable to attacks from birds and other predators.
Having a gill provides a big advantage, as the snail does not have to come to the surface that often. At the surface the apple snail is more vulnerable to attacks from birds and other predators.
Unfortunately, the gill alone does not provide enough oxygen at all times (see above), but apple snails have developed a solution for this: the siphon. With such siphon (a breathing tube formed by a fold of the left mantle cavity) apple snail can breathe air while staying hidden below the surface.
Description of the respiratory organs:
The ctenidium is the gill of the apple snail.
This organ can be found attached at the roof of the right mantle cavity (the branchial chamber), starting close at the edge of the mantle cavity and ending deep inside the right mantle cavity. From the outside of thin-shelled snails it can be seen as a slightly darker strip, next to the lung.
The ctenidium consists of a row of gill filaments that hang into the mantle cavity. These filaments considerably enlarge the exchange surface for oxygen and carbondioxide (CO2). To obtain a efficient respiration with the gill, the water in the mantle cavity has to be refreshed constantly: water enters at the left side (where the inhalend siphon is situated), flows over the osphradius (smell organ), then over the gill and the renal aperture (urine opening) and then leaves the mantle cavity trough the exhalend siphon, after passing the anus. This constant water flow in the mantlecavity is created by the cilia on the surface of the snail skin.
The ctenidium is the gill of the apple snail.
This organ can be found attached at the roof of the right mantle cavity (the branchial chamber), starting close at the edge of the mantle cavity and ending deep inside the right mantle cavity. From the outside of thin-shelled snails it can be seen as a slightly darker strip, next to the lung.
The ctenidium consists of a row of gill filaments that hang into the mantle cavity. These filaments considerably enlarge the exchange surface for oxygen and carbondioxide (CO2). To obtain a efficient respiration with the gill, the water in the mantle cavity has to be refreshed constantly: water enters at the left side (where the inhalend siphon is situated), flows over the osphradius (smell organ), then over the gill and the renal aperture (urine opening) and then leaves the mantle cavity trough the exhalend siphon, after passing the anus. This constant water flow in the mantlecavity is created by the cilia on the surface of the snail skin.
 The exhalend siphon at the right side of the head. This siphon is merely a fold of skin instead of a real tube. (Pomacea canaliculata) |  A close up of the ctenidium (gill) in an Asolene (Pomella) megastoma) snail. The thiny filaments, that enlarge the exchange surface considerably, are recognizable. |
 In this apple snail (Asolene (Pomella) megastoma): 1. Pneumostome or lung opening 2. The lung itself. |  A close up of the lung in anAsolene (Pomella) megastoma) snail. The blood-vessel rich wall is readily visible in the photograph. |
The lung of the apple snail is an air filled sac in the left mantle cavity.
The walls of this pulmonary sac are very well supplied with blood to exchange oxygen and carbondioxide (CO2) between the blood and the air in the lung.
In the picture above, one can see the numberous vessels in the wall of the lung.
The lung can be closed of with a sphincter when the snail is submerged. When the snail leaves the water, the lung opening (pneumostome or lung aperture) is held open most of the time to allow a constant airflow in the lung.
Refreshment of the air inside the lung (respiration) when the snail is under water, is achieved by moving the head in and out the shell, while the siphon makes contact with the surface. In this way, the lung expands and contracts to move the air in and out the lung.
The number of body contractions (and thus respirations) varies from 10 to 18 and the duration of each contraction is approximately 1 second.
Take a look at this mpeg movie: [snail_respirate] (199kb) to see an apple snail (Pomacea canaliculata) heading to the surface and making contact with the siphon to breathe (The movie plays at 3 times real time).
The walls of this pulmonary sac are very well supplied with blood to exchange oxygen and carbondioxide (CO2) between the blood and the air in the lung.
In the picture above, one can see the numberous vessels in the wall of the lung.
The lung can be closed of with a sphincter when the snail is submerged. When the snail leaves the water, the lung opening (pneumostome or lung aperture) is held open most of the time to allow a constant airflow in the lung.
Refreshment of the air inside the lung (respiration) when the snail is under water, is achieved by moving the head in and out the shell, while the siphon makes contact with the surface. In this way, the lung expands and contracts to move the air in and out the lung.
The number of body contractions (and thus respirations) varies from 10 to 18 and the duration of each contraction is approximately 1 second.
Take a look at this mpeg movie: [snail_respirate] (199kb) to see an apple snail (Pomacea canaliculata) heading to the surface and making contact with the siphon to breathe (The movie plays at 3 times real time).
| Movies (MPEG1): - Breathing apple snail (Pomacea canaliculata) (199kb, 3x) |
The siphon is an organ consisting of a fold of the mantle cavity (nuchal lobe) at the left side of the neck above the head of the snail. When not in use, the siphon is a barely visible, but when needed, the muscles in this fold contract and the fold changes into a flexible tube-like structure, which enables the snail to breathe air while staying submerged. The siphon of an apple snail can thus be compared with a snorkel.
The length of the siphon varies a lot between the different species: all snails from the family Pomacea have a very long siphon, when fully stretched it's longer than the snail itself (about 2.5 times), while the other genera of the family Ampullariidae (Afropomus, Asolene, Felipponea, Lanistes, Marisa, Pila and Saulea) have shorter siphons (less than the body length).
The length of the siphon varies a lot between the different species: all snails from the family Pomacea have a very long siphon, when fully stretched it's longer than the snail itself (about 2.5 times), while the other genera of the family Ampullariidae (Afropomus, Asolene, Felipponea, Lanistes, Marisa, Pila and Saulea) have shorter siphons (less than the body length).
 Detail of an extented siphon (Pomacea canaliculata). Click on picture to enlarge. |  Breathing when submerged (Pomacea canaliculata). |
 Often overlooked, but Marisacornuarietis, also has a breathing siphon, although much shorter than in the Pomacea species. |  Asolene pixi breathing with its short siphon. |
Subscribe to:
Comments (Atom)